| Issue |
Natl Sci Open
Volume 2, Number 6, 2023
|
|
|---|---|---|
| Article Number | 20220053 | |
| Number of page(s) | 23 | |
| Section | Life Sciences and Medicine | |
| DOI | https://doi.org/10.1360/nso/20220053 | |
| Published online | 03 July 2023 | |
REVIEW
Dendritic cell maturation in the tumor microenvironment
1
Institute of Cancer Research, Shenzhen Bay Laboratory, Shenzhen 518132, China
2
School of Pharmaceutical Sciences, Tsinghua University, Beijing 100084, China
3
BIOPIC and School of Life Sciences, Peking University, Beijing 100871, China
* Corresponding author (email: This email address is being protected from spambots. You need JavaScript enabled to view it.
)
Received:
1
November
2022
Revised:
13
December
2022
Accepted:
21
December
2022
Abstract
Maturation is essential for dendritic cells (DCs) to function as a bridge between innate and adaptive immunity. The plastic maturation states correspond to the multifaceted roles of DCs in the initiation of protective, tolerogenic and pathogenic immune responses. The formation of a distinct maturation state of DCs involves the complex crosstalk between the intrinsic differences in their ontogeny and the immune environments. Recent studies have defined a universal mature DC population in the tumor microenvironment (TME), but their ontogenesis and functions remain unclear. Here, we summarize recent advances in DC maturation and attempt to dissect the complex functions of mature DCs in the TME. We highlighted that there is a distinct similarity and heterogeneity of mature DCs in the TME, in which the environment and development are both important. These help us revisit the diversity of DC maturation and to better dissect the complex functions of DCs in the TME.
Key words: dendritic cells / tumor microenvironment / tumor-associated mature DCs / tolerogenic DCs / tertiary lymphatic structures / immunotherapy
© The Author(s) 2023. Published by Science Press and EDP Sciences
 This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Introduction
Dendritic cells are professional antigen-presenting cells (APCs). Although conventional DCs (cDCs), plasmacytoid DCs (pDCs) and monocyte-derived DCs (moDCs) are or have been named DCs based on similar morphology or surface markers, only cDCs exhibit the ability to prime naïve T cells alone in vivo [1]. Considering that the primary function of DCs is to initiate adaptive immune responses, we will mainly focus on the maturation of cDCs here and will only introduce features of pDCs or moDCs when illustrating the mechanisms that may be common in cDCs. cDCs are conserved across multiple species [2], suggesting their irreplaceable roles in the adaptive immune responses. There are several developmentally, phenotypically and functionally distinguishable conserved DC subsets in both humans and mice. The development and classification of DCs have been widely discussed [1,3]. Briefly, based on phenotypic and developmental differences, DCs are mainly classified into pDCs and cDCs, which in turn can be further divided into cDC1 and cDC2 subsets [4]. In terms of tissue localization, cDCs can be further divided into lymphoid tissue-resident cDCs and migratory cDCs, both of which contain cDC1s and cDC2s [3]. After being exposed to pathogens in tissues, migratory DCs matured and migrated to adjacent draining lymph nodes, where they can prime antigen-specific T cells. Tissue-resident DCs are often colonized in lymphoid tissues, and they can capture antigens from blood, lymph, or antigens delivered by other migratory APCs.
Maturation of DCs is a conserved program that involves many changes in morphology, phenotype, phagocytosis, secretory characteristics, epigenetic modifications and metabolic characteristics [5]. Here we use the term “maturation” to describe the dramatic alterations in DCs after maturation and refrain from the term “activation” to avoid any misunderstanding of mature DC function [5], since mature DCs can also be tolerant. Immature DCs are incompetent for naïve T cell priming as they rarely express co-stimulatory molecules or produce cytokines [6]. Although they are efficient in phagocytosis, excessive degradation of the phagocytosed antigens and the rapid recycling of major histocompatibility complex (MHC) makes it difficult for them to display MHC-peptide complexes [7]. A typical DC maturation process is induced by the activation of pattern recognition receptors (PRRs) triggered by pathogen-associated molecular patterns (PAMPs) or damage-associated molecular patterns (DAMPs), and is characterized by the up-regulation of MHCs and co-stimulatory molecules, as well as increased production of inflammatory factors, which enable them to prime antigen-specific naïve T cells and to initiate the optimal adaptive immune responses [8]. However, abnormal DC maturation could provoke pathological immune responses leading to autoimmune diseases [9], or graft-versus-host diseases [10].
Multiple studies have suggested that different DC subsets exhibit distinct functions upon maturation, and they might be involved in distinct types of immune responses [11,12]. cDC1s are mainly involved in sensing intracellular bacteria and viruses, mediating the T helper (Th)1 cell polarization of CD4+ T cells and the differentiation of CD8+ T cells into cytotoxic T lymphocytes (CTLs), thereby initiating the type I immune responses. In this process, pDCs serve as an important complement in response to viral infections by producing type I interferons (IFNs). The Klf4+ or Irf4+ cDC2s may be involved in parasite- or allergen-induced Th2 cell polarization and type II immune responses. Similarly, the Notch-dependent cDC2s may be involved in Th17 cell polarization and type III immune responses. These paradigms seem to over-emphasize the determinacy of DC ontogeny on their functions. The extrinsic features such as their spatial distribution in tissues are also important for functional differences [13]. Additionally, in the house dust mite (HDM) infection model, the different maturation states, rather than the developmental differences of cDC2s, determine the Th2 cell and Th17 cell polarization [14]. Furthermore, both cDC1s and cDC2s are non-redundant during the induction of follicular helper T cells and peripheral regulatory T cells (Tregs) [3]. Thus, despite the early viewpoints emphasizing the lineage determinism of DC functions, accumulating evidence suggests that DC functions are also context dependent.
In this review, we summarize the current understanding of the DC maturation to decode the function of mature DCs in the TME. We also review how the maturation characteristics of DCs are reshaped by the TME to achieve immune escape. Finally, we elucidate how tumor therapy remodels the DC maturation.
The features of DC maturation
DC maturation is a stepwise program
Increased evidence suggests that the maturation of DCs is not a simple activation process, but a stepwise program with multiple outcomes, including stimulatory DCs, tolerogenic DCs, hyperactivated DCs and pyroptotic DCs.
For a long time, tolerogenic DCs have been considered as immature or semi-mature DCs. But this hypothesis may be skewed, since recent studies have shown that tolerogenic DCs also underwent a complete maturation program. For example, tolerogenic DCs actively express immunosuppressive co-stimulatory molecules like PD-L1 and PD-L2 and anti-inflammatory factors, such as indoleamine 2,3-dioxigenase 1 (IDO1) [15], in contrast with the quiescent states of immature DCs. In addition, cDC1s mediating central tolerance under steady state in mice exhibit a convergent gene expression alteration compared to toll-like receptor (TLR)-stimulated mature cDC1s [16]. Furthermore, differential expression of hundreds of genes exist between tolerogenic cDC1s and their immature counterparts, indicating that they are not in the immature state [16]. Interestingly, the tolerogenic DCs seem to represent a remodeling of maturation rather than a de novo maturation program. For example, IL-10, IL-27, dexamethasone, minocycline, vitamin D, rapamycin, Transforming growth factor (TGF)-β, cyclosporine, tacrolimus, aspirin, retinoic acid (RA) and the aryl hydrocarbon receptor ligands have been shown to induce tolerogenic DCs under tumor or inflammatory conditions [17], but many of them cannot induce DC maturation alone.
The classical DC maturation may also be regulated by additional remodeling processes. For example, in mouse infection models, mature DCs initiate an effective CTL response that is dependent on “helper” signals delivered by CD4+ T cells [18,19]. These “helper” signals are induced by CD40L, exosomes, and type I IFNs from CD4+ T cells, and upregulate the expression of co-stimulatory molecules and the production of cytokine IL-12 of mature DCs [20,21]. Similarly, in a mouse fibrosarcoma model, the CD40-CD40L interaction between CD4+ T cells and cDC1s is necessary for the initiation of CD8+ T cell responses [22]. In addition, although CD40L alone can induce activation of immature DCs in vitro, immature cDCs from germ-free mice appear to be tolerant for CD40L stimulation [23], suggesting that CD40L is more likely to remodel the mature state of DCs rather than to induce a de novo DC maturation. Furthermore, semi-mature cDC2s and fully mature cDC2s seem to play distinct functions in the HDM infection model [14], where the semi-mature cDC2s promote Th17 cell differentiation, but the fully mature cDC2s induce strong Th2 cell differentiation and suppress Th17 cell differentiation at the same time.
The inflammasome-mediated DC maturation provides additional evidence for this stepwise model. In macrophages and DCs, the activation of inflammasomes usually induces a massive release of active IL-1β, accompanied by a rapid pyroptosis [24]. However, the combination of oxidized lipids and TLR ligands induces DCs into a long-lasting “hyperactivated” state, reflected by the maintenance of cell viability and the production of large amounts of cytokines including IL-1β [25]. In addition, in mouse melanoma models, oxidized lipid-induced hyperactivation of cDC1s and cDC2s significantly enhances their migration and induces long-lasting CTL antitumor immunity [26]. Furthermore, upon hyperactivation, but not TLR ligand-induced maturation, DCs produce large amounts of IL-1β, which enhances the cytotoxicity and long-term antitumor capacity of tumor-infiltrating CD8+ T cells [27,28].
These findings support that the DC maturation program may be not a one-step process, but rather a stepwise program (Figure 1). The maturation signals from inflammatory factors and PRR may only provide the basic requirements, and other signals are needed to complete or remodel the DC maturation. This stepwise maturation program allows DCs to rapidly respond to danger signals and adjust their functions precisely according to environmental conditions. Although limited information is known about the mechanisms, it is no doubt that they are important for mature DCs to perform diverse functions.
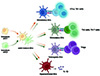 |
Figure 1 The stepwise model of DC maturation. The semi-mature DCs are “ready” to receive further stimulations to become fully-mature DCs and to perform diverse functions. |
DC maturation reflects an integration of stimulatory signals
The stepwise maturation model brings diversity to mature DCs but does not lead to an infinite type of mature DCs. Indeed, despite the exposure to an unpredictable number of stimuli and stimulus combinations, the intrinsic features of DCs enable the integration of input signals, so that DCs only exhibit several conserved maturation states. For example, upon lipopolysaccharides-induced bone marrow-derived DC maturation in vitro, Perturb-seq has revealed the existence of four transcription factor regulatory modules in DCs that collectively regulate five programs of DC maturation, suggesting that the existence of a common pattern of DC maturation [29]. Similarly, human moDCs exhibit five convergent conserved maturation states in response to different stimuli [30]. These states are correlated with different Th cell polarization, suggesting that maturation states of DCs determine their preference to initiate adaptive immune responses [30]. Additionally, two conserved modules of mature cDC2s have been found across cancer and other diseases: the secretory type with PD-L1hi ICOSLlow and the helper type with PD-L1low ICOSLhi [31]. Furthermore, mature migratory cDC2s express similar co-stimulatory molecular signatures in response to different types of pathogen infection, and the main driver for Th cell polarization lies in different cytokine profiles [32].
Therefore, although DC maturation is a stepwise program with multiple outcomes, only several conserved maturation states are observed, which may be attributed to the integration of diverse stimulus signals by several intrinsic regulatory modules of DCs. This also allows mature DCs to function in an organized rather than a chaotic manner in different contexts.
Functions of tumor-associated mature DCs
Maturation is critical for DC-mediated tumor control
Although mature DCs can be immunostimulatory or immunosuppressive, many studies still emphasize the importance of DC, especially DC maturation, for tumor control [33–35]. For cDC1s, in mouse melanoma models, deficiency of IKKβ (downstream of NF-κB) or IRF1 signaling (downstream of IFN-γ) in cDC1s impairs their maturation and the initiation of CD8+ T cell responses [36]. Similarly, in melanoma patients, a cDC1 signature containing genes of NF-κB and IRF1 pathway is associated with improved prognosis compared with the cDC1 signature alone [36]. For cDC2s, early studies have shown that cDC2s are associated with type II immune response, type III immune response or Treg induction in the TME, which may indicate a poor prognosis. However, mature migratory cDC2s also transport tumor antigens to lymph nodes and contribute to the priming of tumor-reactive T cells [37]. Consistently, in a cDC1-deficient fibrosarcoma model, a group of interferon-stimulated gene (ISG)-expressing mature cDC2s facilitates an effective CTL response and tumor control [38].
Although the high expression of cDC signature genes is associated with better prognosis in many mouse models and cancer patients [34,39], recent studies further emphasize that mature cDC signature genes, compared with the general cDC signature genes, are better anti-tumor and prognostic biomarkers. For example, in melanoma patients, there is a strong correlation between Trms and mature DCs, but a weaker correlation with immature DCs [40]. In addition, the proportion and maturation state of migratory cDCs is negatively correlated with primary tumor size [41]. Similarly, in mouse melanoma models, extracellular vesicles from mature DCs, rather than those form immature DCs, are more efficient to induce antigen-specific CD8+ T cell activation [42]. Furthermore, in human neuroblastoma, both DC signature genes and natural killer (NK) cell signature genes are positively correlated with T cell infiltration at the transcriptome and protein levels and are associated with a good prognosis [43]. Notably, DC signature genes used in this study, such as CCL17 and CCL22, are highly expressed in mature cDCs specifically. Likewise, in gastric cancer, the TME features with high expression of mature DC signature genes are also better for prognosis than the TME features containing immature DCs [44].
Therefore, these studies suggest that the maturation status rather than the developmental features of DCs determine their roles in the TME, although cDC1 seems to play more important roles in tumor control compared to other DC subsets. However, the DC maturation features described in these studies are more similar to those of stimulatory DCs, whether tolerogenic DCs, hyperactivated DCs and pyroptotic DCs also contribute to tumor control is still unclear.
PRR activation contributes to tumor control
In line with the essential roles of stimulatory DCs in tumor control, the activation of the cyclic GMP-AMP synthase-stimulator of interferon genes (cGAS-STING) pathway and TLRs also play important roles in tumor control, as both of them are classical inducer of stimulatory DCs. For example, DNA from tumor cells, or microbes within the tumor, trigger the activation of the cGAS-STING pathway in cDC1s, inducing cDC1 maturation and production of type I IFNs [45–47], which is required for effective cross-presentation and tumor-specific CTL priming. In addition, induction of DC maturation with TLR or STING agonists is associated with type I IFN production and CD8+ T cell-mediated anti-tumor efficacy [48]. Furthermore, STING agonists also indirectly activate NK cells in an IL-15/IL-15Ra-dependent manner by promoting IL-15Ra expression on DCs [49]. Immunotherapies also cause DC maturation via the cGAS-STING pathway. CD47 blockade restricts the degradation of tumor mitochondrial DNA in DCs but not in macrophages in mouse tumor models, thereby promoting the activation of the cGAS-STING pathway in DCs, which promotes the type I IFN production and induces effective CTL responses [50]. Similarly, Tim-3 blockade promotes phagocytosis of extracellular DNA by cDC1s, and the phagocytosed DNA activates the cGAS-STING pathway, which in turn promotes the production of CXCL9 and type I IFNs by cDC1s [51]. Together, all these studies emphasize the importance of cGAS-STING pathway-triggered DC maturation and IFN release for tumor control.
TLRs are the most widely known PRRs that trigger stimulatory maturation of DCs. Multiple studies have highlighted a clear correlation between the antitumor effects of TLR agonists and their induction of DC maturation. Although most tumor-infiltrating DCs exhibit an immature phenotype, in vitro treatment with TLR8 agonists in combination with IFN-γ or poly I:C effectively induces their maturation, suggesting the feasibility of therapeutic interference to modify the maturation state of DCs [52]. Consistently, the induction of cDC1 maturation by TLR3 agonists in breast cancer leads to the production of type III interferon (also known as IFN-λ), which facilitates Th1 cell polarization and CD8+ T cell infiltration [53]. Similarly, intratumoral injection of TLR9 agonists induces DC maturation and improves the responses to PD-1 blockade therapy in melanoma patients [54]. Furthermore, TLRs agonists, when combined with Fms-related tyrosine kinase 3 ligand (Flt3L), induce the expansion of DC progenitors and DC maturation in mouse melanoma, thus enhancing the tumor response to BRAF and immune checkpoint blockade [55]. Likewise, an in situ vaccine that combines Flt3L, radiotherapy and TLR3 agonist induces anti-tumor CD8+ T cell responses and systemic cancer remission in patients with advanced inert non-Hodgkin’s lymphoma [56]. Although TLR agonists provoke excessive immune-related side effects during treatment, there is no doubt that their therapeutic effects are directly linked to the induction of DC maturation.
Therefore, these studies further emphasize the essential roles of stimulatory mature DCs in tumor control and the induction of them is a promising immunotherapeutic strategy.
LAMP3+ DCs are conserved in a variety of tumors
Single-cell transcriptomic studies have provided new insights into the maturation status of DCs in the TME. Recent studies have defined a conserved mature DC population named LAMP3+ DCs [57] (also known as mregDCs [58], DC3s in some studies [59,60]) in multiple types of cancers and other diseases (Table 1) [31,57–59,61–92]. Here we avoid using the name DC3 to prevent potential confusion with another newly identified DC population, which has a distinct ontogeny and has both cDC2 and monocyte characteristics [93,94]. We use LAMP3+ DCs instead of mregDCs, because the function of this group of cells may not necessarily be immunosuppressive, and we will discuss the diverse and paradoxical functions of these cells later on.
Human LAMP3+ DCs described in the single cell transcriptomics of health and diseases
Although LAMP3+ DCs are conserved across cancer types [61], they do not appear to represent a novel DC subset that developed independently. Firstly, there is still no experimental evidence describing their direct progenitors or precursors. Whether their development is also dependent on Flt3L or Granulocyte-macrophage colony-stimulating factor (GM-CSF) is also unclear. Secondly, transcriptomic data show that they share similarities with both cDC1s and cDC2s, suggesting that they may represent a convergent maturation state of cDC1s and cDC2s, both of which downregulate the expression of their signature genes upon maturation [4]. It is more likely that LAMP3+ DCs may represent a conserved convergent maturation state of different cDC subsets across cancer types.
Considering the importance of DC maturation for tumor control, LAMP3+ DCs which have typical maturation features may be involved in the anti-tumor immune responses. Indeed, LAMP3+ DCs, at least a fraction of them, appear to induce CTL responses. For example, in mouse melanoma models, migratory mature cDCs occupy a perivascular niche, produce CXCL16, and recruit CXCR6+ CD8+ T cell aggregates [95]. They also express IL-15/IL-15RA, which acts on adjacent CXCR6+ CD8+ T cells to promote their proliferation and survival [95].
However, the most important difference between LAMP3+ DCs and the mature stimulatory DCs is that they also highly express immunoregulatory factors such as IDO1 and IL4I1 [57], thus multiple studies believe that LAMP3+ DCs may be favoring a tolerogenic function [58]. Indeed, compared with other tumor-infiltrating immature DCs, LAMP3+ DCs highly express CCL17 and CCL22, which may promote the recruitment of CCR4+ Tregs to the TME [84]. In addition, in gliomas, LAMP3+ DCs exhibit immune tolerance characteristics such as reduced antigen presentation and increased expression of immunomodulatory molecules [96].
The third notable feature of LAMP3+ DCs is their high expression of immune checkpoints such as PD-L1 and PD-L2, suggesting their possible contribution to the induction of immune exhaustion. In fact, the expression of immune checkpoints may be a negative feedback mechanism that appears along with DC maturation to prevent excessive immune responses or damage of DCs by cytotoxic cells [97]. In addition, even if LAMP3+ DCs are involved in the induction and maintenance of exhausted T cells, it does not necessarily mean a disruption of anti-tumor immunity, because the existence of exhausted T cells in the context of chronic inflammation is often associated with long-lasting anti-viral and anti-tumor effects. For example, cDC1s maintain exhausted CD8+ T cell precursor niches during chronic lymphocytic choriomeningitis virus infection, and these niches ensure the existence of long-lasting anti-viral immunity and allow the expansion of the exhausted T cell precursor during PD-L1 blockade [98]. Similarly, in mouse lung adenocarcinoma models, cDC1s in tumor-draining lymph nodes maintain the existence of a TCF1+ CD8+ T cell pool (precursor of exhausted T cells), which is essential for effective anti-tumor immunity [99]. Furthermore, the abundance of cDC1s in tumor-draining lymph nodes decreases with the progression of mouse lung adenocarcinoma, and Flt3L combined with CD40 agonists restores cDC1 abundance and the expression of co-stimulatory molecule in tumor-draining lymph nodes, enhancing CD8+ T cell responses and tumor control [99].
The combination of immunostimulatory and immunomodulatory features makes LAMP3+ DCs difficult to be classified as a well-established DC maturation type. However, they may also not represent a tumor-specific DC maturation state, as LAMP3+ DCs have also been found in health and other diseases. A more favorable possibility is that LAMP3+ DCs represent a collection of DCs in different maturation states in the TME, distinguished from other immature DCs due to the huge transcriptome alterations upon maturation. In addition, the low abundance of LAMP3+ DCs in the tumor microenvironment makes it difficult to capture enough cell numbers to further analyze their heterogeneity. Consistently, a pan-cancer single-cell transcriptional analysis shows that LAMP3+ DCs can be further divided into two functional heterogeneous clusters, where cDC1-derived LAMP3+ DCs predominantly receive IL-12 and IL-4 signaling and overexpress IL12B (inducing Th1 polarization) [68], while cDC2-derived LAMP3+ DCs predominantly receive IFN family signaling and overexpress CCL17, recruiting CCR4+ Tregs (Figure 2A) [65]. This also suggests a preference in the TME where cDC1s may prefer to enter the stimulatory maturation state, while cDC2 may contribute to the main source of tolerogenic DCs.
 |
Figure 2 The potential functional heterogeneity of LAMP3+ DCs. (A) cDC1-deriverd LAMP3+ DCs mainly produce CXCL16 and IL-15, which promote the recruitment and maintenance of CD8 T cells in the TME and therefore enhance the antitumor immune effects. cDC2-deriverd LAMP3+ DCs highly express CCL17, CCL22 and IDO1, which promote the recruitment and differentiation of Treg in the TME and suppress the antitumor immune response. (B) LAMP3+ DCs within TLSs actively recruit and interact with T cells to elicit neoantigen-specific T cell activation, or Treg production. LAMP3+ DCs outside of TLSs exhibit impaired antigen-presenting ability and may be involved in immunosuppression. |
Thus, emerging evidence from single-cell transcriptomics suggests the existence of multiple maturation states of DCs in the TME. Also, the newly identified LAMP3+ DCs may represent a collection of DCs in different mature states that have similar transcriptional profiles but distinct functions. A more detailed mapping of the maturation states of LAMP3+ DCs would help understand the relationship between tumor-associated mature DCs and well-known DC maturation states such as stimulatory DCs, tolerogenic DCs, hyperactivated DCs and pyroptotic DCs. This will also help us to understand the unique mechanisms of DC maturation in the TME, as well as to better operate the DC maturation for therapy.
LAMP3+ DCs affect the formation and maintenance of TLSs
TLS is a lymphocyte aggregate without a pericyte membrane that is typically found in chronic inflammatory conditions and tumors [100]. Similar to secondary lymphoid organs, TLSs also have T cell-rich regions, high endothelial venule (HEV) structures, as well as germinal center structures in mature TLSs [100]. Mature DCs are essential components of the TLSs. The marker of mature DCs, LAMP3, is often used alone or together with CD20 to define TLSs [100,101]. Consistently, in multiple types of cancers, LAMP3+ DCs tend to be enriched in TLSs [101], especially in non-small-cell lung cancer (NSCLC), where LAMP3+ DCs almost exclusively appear in the TLSs. However, in HCC and clear cell renal carcinoma (ccRCC), mature DCs also exist outside TLSs [102–104].
Mature DCs play an indispensable role in the formation of TLSs. In human squamous cell carcinoma, mature migratory DCs display the highest CCL19 and CCL21 expression, both of which are critical for naïve T cell homing and TLS formation [64]. Consistently, a pan-cancer analysis also supports that LAMP3+ DCs express higher levels of CCL19 than other immature DCs in multiple types of cancers [60], in line with their enrichment in TLSs. In addition, treatments that induce DC maturation also promote the TLS formation. For example, CD40 agonists induce the maturation and production of LTα, LTβ and LIGHT of DCs, facilitating the formation of TLS-like structures and the aggregation of T cells in gliomas [105]. Similarly, low-dose STING agonist treatment of mouse melanoma also induces the maturation and production of TLS-inducing cytokines, such as LTα, IL-36β and TNF-α, by DCs [106].
Additionally, LAMP3+ DCs are essential for the initiation of tumor-specific adaptive immune responses in TLSs. The tumor-specific adaptive immune responses initiated by LAMP3+ DCs in TLSs may represent the major paradigm of antitumor immunity, as the presence of TLSs is associated with better responsiveness to immune checkpoint blockade (ICB) therapies, both in tumors with high tumor mutational burden (TMB), such as NSCLC and melanoma, and in tumors with low TMB, such as soft tissue sarcoma and ccRCC [107]. In addition, LAMP3+ DCs in the germinal center trigger more efficient antigen presentation in an IgG-dependent manner [108]. Furthermore, in human breast cancer, the density of LAMP3+ DCs is tightly correlated with the abundance of HEVs, likely attributed to the LTβ production by LAMP3+ DCs [109]. A notable exception is ccRCC, where 80% of LAMP3+ DCs are located outside of the TLS and do not express co-stimulatory molecules such as CD83 and show low expression of MHC-Ⅱ [100]. Interestingly, high non-TLS DC density is a hallmark of advanced TNM staging and predicts a worse prognosis [110]. Consistently, the LAMP3+ DC density is associated with shorter survival in ccRCC with lung metastases [100]. These results suggest that LAMP3+ DCs within and outside TLSs may be in different maturation states, allowing them to perform different functions (Figure 2B).
Abnormal maturation of DCs in the TME
The non-immune component of the TME inhibits DC maturation
In the TME, DCs exhibit not only impaired differentiation and tumor infiltration [111–114], but also abnormal maturation [34,115]. The abnormal maturation of DCs seems to be actively regulated by multiple components in the TME. Prostaglandin E2 (PGE2) produced by tumor cells not only impairs the recruitment of DCs via tumor-infiltrating NK cells, but also directly inhibits DC maturation [116]. In vitro, PGE2 inhibits DC maturation by suppressing the recognition of DAMPs on DCs [35]. Consistently, gemcitabine treatment triggers the typical immunostimulatory DAMP releases (e.g., calreticulin, HSP70 and HMGB1) but fails to induce immunogenic cell death (ICD), in large part due to the parallel triggering of PGE2 release, which inhibits cDC1 maturation [117]. In addition, aberrant epigenetic modifications in tumor cells, such as increased m6A modification of mRNA, affect the MHC expression and IL-12 production during DC maturation [118]. Consistently, increased m6A modification of mRNA in tumor-infiltrating DCs also leads to increased translation of lysosomal proteases, resulting in excessive antigen degradation and thus inhibition of antigen presentation [119]. Furthermore, vascular endothelial growth factor (VEGF) [35,120] and α-fetoprotein [121] expressed by tumor cells also inhibit the maturation of tumor-infiltrating DCs, thus promoting tumor immune escape. All these studies emphasize that the intrinsic properties of tumor cells inhibit the maturation of tumor infiltrating DCs and the immunostimulatory capacity of mature DCs.
The importance of metabolic shifts for DC maturation has been extensively discussed [115,122]. Briefly, DC maturation is accompanied by a shift in the energy supply manner to fulfill the energetic and biosynthetic requirements during DC maturation. Tumor-infiltrating DCs exhibit aberrant metabolic paradigms, which in turn impair their maturation program and remodel their mature phenotype and function. Hypoxia [123–125], ER stress [126], nutrient deficiency [115], accumulation of reactive oxygen species [126], aberrant amino acid metabolism (tryptophan, arginine, etc.) [127], and aberrant adenosine metabolism [128] in the TME show a marked inhibitory effect on the expression of co-stimulatory molecules and migration during DC maturation. In addition, these also induce the production of anti-inflammatory factors such as IL-10 and reduce the production of pro-inflammatory factors such as IL-12 of mature DCs. Furthermore, in contrast to DC hyperactivation induced by oxidized lipids, lipid accumulation in tumor-infiltrating DCs impairs their antigen processing and presentation [35,129–131].
In addition to tumor cells and the TME metabolic profiles, other components of the TME also affect DC maturation. For example, vitamin E enhances the cross-presentation of DC-derived extracellular vesicles, triggering systemic antigen-specific T cell antitumor immunity [132]. Oral vitamin E improves the survival of cancer patients receiving ICB therapies [132]. In addition, short-chain fatty acids produced by intestinal microbes interfere with the IL-12 production of mouse LAMP3+ DCs, thereby impairing radiotherapy-induced CD8+ T cell responses [133]. Furthermore, in human and mouse squamous cell carcinomas, WNT2 expressed by cancer-associated fibroblasts (CAFs) negatively correlates with DC maturation and CD8+ T cell activation [134]. Anti-WNT2 antibody restores anti-tumor T cell responses and enhances anti-PD-1 efficacy by increasing DC maturation. Moreover, IL-1α and IL-1β in the TME enhance thymic stromal lymphopoietin (TSLP) secretion from CAFs, and TSLP promotes the polarization of CD4+ T cells to Th2 cells by increasing the expression of OX40L, CD80 and CD86 on DCs, as well as chemokine production [135].
In addition to inhibiting and remodeling the DC maturation, the TME also affects the survival and migration of mature DCs. For example, the decreased abundance of cDCs in the sentinel lymph nodes of melanoma is attributed to their excessive phagocytosis-induced apoptosis induced by melanoma-secreted lysosomes [136]. Additionally, tumor-derived liver X receptor (LXR) ligands impair the expression of CCR7 on DCs and disrupt the migration of DCs to tumor-draining lymph nodes [137]. Furthermore, tumor-derived factors inhibit the expression of CCL21 in stromal cells, which is required for the recruitment of T cells and DCs [137]. Collectively, these studies suggest that many features of tumor cells and the TME directly regulate DC maturation to remodel tumor immunity (Figure 3).
 |
Figure 3 Mechanisms regulating the tumor-infiltrating DC maturation. Tumor cells, CAFs, and tumor-infiltrating microbes inhibit the maturation of tumor-infiltrating DCs by regulating metabolism, etc. The effector T cells and NK cells in the tumor microenvironment promote the generation of LAMP3+ DCs by producing chemokines and IFNs, and immunosuppressive cells such as Tregs inhibit the generation of LAMP3+ DCs by producing IL-10 and TGF-β. The efficacy of radiotherapy and other treatments also depends on the induction of LAMP3+ DCs. The efficacy of radiotherapy, chemotherapy and immunotherapy is also dependent on their induction of LAMP3+ DCs. |
Immune cells in the TME regulate DC maturation
Both tumor-infiltrating NK cells and T cells appear to be associated with DC recruitment and maturation. The necessity of “help” signaling delivered by CD4+ T cells for DCs to initiate CTL responses has been described above. NK cells also affect DC maturation in the TME. For example, in mouse breast cancer models, CCL5 and XCL1 secreted by NK cells recruit cDC1s to the TME, enhancing tumor rejection and immunotherapy response [116]. PGE2 impairs chemokine and FLT3L production of NK cells, thereby inhibiting the infiltration and survival of cDC1s in the TME [35,116]. Consistently, in human melanoma, the abundance of stimulatory DCs is correlated with NK cell abundance and FLT3LG levels [138].
The crosstalk between Tregs and DCs has also been discussed [3]. On the one hand, tolerogenic DCs in the TME facilitate Treg differentiation by producing anti-inflammatory factors and increase Treg accumulation by producing chemokines such as CCL17 and CCL22. On the other hand, Tregs also actively remodel the characteristics of tumor infiltrating DCs. For example, in human and mouse melanoma, the balance between Tregs and cDC2s determines the activation of T cells and prognosis [139]. Treg depletion results in elevated expression of co-stimulatory molecules on mature cDC2s in the tumor-draining lymph nodes and promotes the differentiation of effector T cells [139]. Similarly, in mouse orthotopic pancreatic ductal adenocarcinoma models, Treg depletion also boosts the accumulation of intratumoral DCs and their expression of co-stimulatory molecules [130]. The inhibitory effect of Tregs on DC maturation is largely attributed to the production of IL-10 and TGF-β [120,140], but direct contact between Tregs and DCs also plays non-redundant roles. For example, in mouse models, CTLA-4 on Tregs decreases the expression of co-stimulatory molecules of mature cDCs, leading to looser conjugation between cDCs and Th cells, thus preventing Th cell activation [141]. Similarly, classical Tregs have been demonstrated to directly interact with LAMP3+ DCs and suppress their immunogenic function by inhibiting the expression of co-stimulatory molecules required for CD8+ T cell activation [142].
Altogether, these results suggest that DC maturation is inhibited and remodeled by multiple components in the TME (Figure 3). Nevertheless, a certain number of LAMP3+ DCs with mature features still exist in multiple kinds of tumors. They may perform distinct functions, even a fraction of them is anti-tumorigenic. However, in most cases, the insufficient abundance prevents them from triggering an effective anti-tumor immune response, probably resulting in cold tumors. Even if LAMP3+ DCs are capable of provoking robust immune responses, the subsequent T cell exhaustion further leads to a failure of tumor control, resulting in hot tumors.
Remodeling of DC maturation by various therapies
Mature DCs are involved in the response to immunotherapy
DC maturation seems to play an important role in a variety of different therapies, including those by CD40 agonists, ICB and phagocytic checkpoint blockade. Since CD40L signaling is important for the DC function to initiate tumor specific CD8+ T cell responses, it has been expected that the use of CD40 agonists can effectively trigger or remodel the maturation state of tumor infiltrating DCs [143]. In mouse bladder cancer models, tumor-infiltrating DCs highly express CD40 and treatment with CD40 agonists elicits an effective anti-tumor CD8+ T cell response and tumor regression [144]. Similarly, in mouse pancreatic ductal adenocarcinoma models, the presence of cDC1s is essential for CD40 agonists to trigger and maintain the anti-tumor CD8+ T cells [145]. Additionally, in mouse colorectal cancer models, CD40 agonist treatment enhances cDC1 maturation (with typical LAMP3+ DC signatures) and increases the abundance of Bhlhe40+ Th1-like cells and CD8+ memory T cells [80].
Recent studies have shown that PD-1/PD-L1 interaction in tumor-draining lymph nodes, but not in tumors, is correlated with better prognosis in multiple types of tumor-bearing mice. In primary tumors and tumor-draining lymph nodes, DCs express the highest levels of PD-L1 [146,147], and blockade of PD-L1 on DCs, but not on macrophages, elicits effective anti-tumor immunity [97,146,148]. Additionally, PD-L1 blockade not only affects the trans PD-L1: PD-1 interaction between DC-T cells, but also affects the cis PD-L1: CD80 interaction on DCs [149]. The disruption of the cis interaction between PD-L1 and CD80 accelerates tumor growth [150], while disruption of the trans interaction between PD-L1 and PD-1 effectively releases CD80 and enhances T cell activation [151]. Notably, LAMP3+ DCs express the highest PD-L1 and are predominantly localized in TLSs and tumor-draining lymph nodes, but whether LAMP3+ DCs are critical for the response to immune checkpoint therapy remains to be investigated.
In addition to PD-L1, PD-1 and Tim-3 blockade may also affect DC maturation. Breast cancer patients treated with PD-1 have been found to have increased numbers of LAMP3+ DCs, suggesting an active anti-tumor immune response [82]. Similarly, in mouse model, PD-1 treatment-induced IFN-γ promotes cDC1 maturation, which further enhances the CD8+ T cell activation [152]. In addition, CXCL9 produced by CD103+ DCs is required for the anti-PD-1 responses by regulating the interaction between CD8+ T cells and DCs [153]. Furthermore, CTLA-4 and PD-1 blockade increases the production of IL-33, which acts directly on DCs to increase their number and maturation, thereby enhancing CD8+ T cell responses [154]. These studies suggest that PD-1 blockade may directly or indirectly enhance the maturation of DCs. Similarly, Tim-3 is highly expressed in tumor-infiltrating CD103+ DCs in spontaneous breast cancer mice [155]. Tim-3 blockade during chemotherapy enhances CXCL9 expression on cDC1s, thereby enhancing CD8+ T cell responses [156]. Tim-3 blockade also promotes phagocytosis of extracellular DNA by DCs, which in turn promotes CXCL9 and type I interferon production by cDC1s [51].
Phagocytic checkpoint blockade, specifically the CD47-SIRPα axis, is associated with the alteration of DC maturation [157,158]. DCs in human and mouse HCC highly express SIRPα, and silencing SIRPα expression induces higher IL-12 production and co-stimulatory molecule expression, leading to a more robust cytotoxic T lymphocyte response [159]. Consistently, in mouse HCC models, CD47 blockade induces activation of the cGAS-STING pathway in DCs, facilitating the production of IL-12 and CXCL9, which promote the recruitment and activation of NK cells [160]. In particular, CD47 blockade restricts the degradation of tumor mitochondrial DNA in DCs but not in macrophages, and tumor mitochondrial DNA activates cGAS-STING pathway in DCs, promoting type I IFN production and CTL effector functions [50]. However, unlike in mice, blocking CD47 impairs both SIRPα and SIRPγ binding without altering T cell responses [161]. This may be due to a greater disagreement between the function of human SIRPγ and that in mice, whereas selective human SIRPα antagonistic antibodies effectively promote chemokine production by macrophages and enhance the cross-presentation capacity of DCs [161].
The maturation of DCs is critical for the ICD effect induced by radiotherapy and chemotherapy
The ICD effect of radiotherapy is critical for long-lasting and distal tumor control [162]. The stress and death of cancer cells during cancer treatment lead to the release of DAMPs, including ANXA1, CALR and HMGB1, which promote the recruitment and maturation of DCs and help initiate effective CTL effector functions. Recent studies have found that DCs not only survive in irradiation, but also sense the DNA and exosomes released by irradiated tumor cells, thereby inducing DC maturation and type I IFN production [163]. These may be attributed to the activation of the cGAS-STING pathway as well as the RIG-I-like receptor LGP2 in DCs by irradiated tumor cells [120,164]. In addition, the combined therapy of low-dose radiotherapy (LDRT), low-dose cyclophosphamide and immune checkpoint blockade mediates the reprogramming of DC maturation in mouse orthotopic ovarian cancers [165]. This reprogramming of DC maturation allows them to become excellent T cell boosters. Likewise, the combination of radiotherapy and IL-15 treatment recruits and induces the maturation of cDC1s, which are critical for the induction of CD8+ T cell-mediated tumor regression and long-term protective memory responses in mouse models [166].
Similarly, certain chemotherapeutic drugs provoke ICD and enhance DC maturation. For example, the telomere targeting drug 6-thio-dG induces DC maturation via the cGAS-STING pathway, thus promoting anti-tumor CD8+ T cell responses [167]. In contrast, in patients and mouse melanoma models, MAPK inhibitors lead to a decreased number of tumor infiltration of cDC1s upon relapse, resulting in cross-resistance to immunotherapy [168]. This may be attributed to the restricted maturation of cDC1s, as the MAPK pathway is also critical for DC maturation. Furthermore, 5-fluorouracil and oxaliplatin directly induce DC maturation, characterized by increased secretion of IL-12 and reduced expression of immunosuppressive ligands, including PD-L1 [169].
Overall, these studies suggest that the immune consequence of traditional therapeutic strategies such as radiotherapy and chemotherapy is associated with the induction or remodeling of the maturation state of LAMP3+ DCs (Figure 3). The maturation state of DCs provoked by therapy is often immunostimulatory and contributes to the establishment of effective CD8+ T cell responses.
Concluding remarks
In this review, we summarize the recent advances in DC maturation. We propose that DC maturation may not be a one-step process, but a stepwise program under strict regulation. The maturation state of DCs is determined by their ontogeny and environmental features and decides their induction of immune stimulation, immune exhaustion or immune tolerance. The reliability and detailed mechanisms of this stepwise model remain to be further investigated. Additionally, how cDCs develop a finite number of conserved maturation states under health and diseases, and what functions these states correspond to remain opaque.
In the TME, the maturation state of DCs is also open to debate. LAMP3+ DCs have been identified in multiple studies using single cell transcriptome sequencing. LAMP3+ DCs are conserved in different cancers and display a unique maturation state characterized by the high expression of immunomodulatory molecules. We emphasize that LAMP3+ DCs are heterogeneous in ontogeny and arise from convergent maturation of cDC1s and cDC2s in the TME. The homogeneous transcriptomic profiles do not mask their functional heterogeneity, although the association of this functional heterogeneity with ontogeny or environment remains to be elucidated. Further studies to compare the transcriptomic heterogeneity within LAMP3+ DCs would help to unravel the basis for this functional difference.
The induction of DC maturation and their distinct maturation states are actively regulated by the TME and therapeutic interventions. Elucidating the formation and specific functions of the distinct DC maturation states will allow for the development of improved immunotherapeutic strategies to provoke optimal anti-tumor immune responses.
Funding
This work was supported by the Major Program of Shenzhen Bay Laboratory (S201101004), the Open Program of Shenzhen Bay Laboratory (SZBL2020090501005), the National Natural Science Foundation of China (81988101, 31991171, 91959000), and the Beijing Municipal Science and Technology Commission (Z211100003321005).
Author contributions
Y.M. and Z.Z. designed the work. Y.M. and Y.L. wrote the manuscript with the help of H.T. and Z.Z. All authors have read and improved the manuscript. Z.Z. is a founder of Analytical Biosciences.
Conflict of interest
The authors declare no conflict of interest.
References
- Cabeza-Cabrerizo M, Cardoso A, Minutti CM, et al. Dendritic cells revisited. Annu Rev Immunol 2021; 39: 131-166. [Article] [CrossRef] [PubMed] [Google Scholar]
- Anderson Iii DA, Dutertre CA, Ginhoux F, et al. Genetic models of human and mouse dendritic cell development and function. Nat Rev Immunol 2021; 21: 101-115. [Article] [CrossRef] [PubMed] [Google Scholar]
- Yin X, Chen S, Eisenbarth SC. Dendritic cell regulation of T helper cells. Annu Rev Immunol 2021; 39: 759-790. [Article] [CrossRef] [PubMed] [Google Scholar]
- Villar J, Segura E. Decoding the heterogeneity of human dendritic cell subsets. Trends Immunol 2020; 41: 1062-1071. [Article] [CrossRef] [PubMed] [Google Scholar]
- Steinman RM. Decisions about dendritic cells: Past, present, and future. Annu Rev Immunol 2012; 30: 1-22. [Article] [CrossRef] [PubMed] [Google Scholar]
- Hammer GE, Ma A. Molecular control of steady-state dendritic cell maturation and immune homeostasis. Annu Rev Immunol 2013; 31: 743-791. [Article] [CrossRef] [PubMed] [Google Scholar]
- Roche PA, Furuta K. The ins and outs of MHC class II-mediated antigen processing and presentation. Nat Rev Immunol 2015; 15: 203-216. [Article] [CrossRef] [PubMed] [Google Scholar]
- Hilligan KL, Ronchese F. Antigen presentation by dendritic cells and their instruction of CD4+ T helper cell responses. Cell Mol Immunol 2020; 17: 587–599 [CrossRef] [PubMed] [Google Scholar]
- Ganguly D, Haak S, Sisirak V, et al. The role of dendritic cells in autoimmunity. Nat Rev Immunol 2013; 13: 566-577. [Article] [CrossRef] [PubMed] [Google Scholar]
- Zhang L, Chu J, Yu J, et al. Cellular and molecular mechanisms in graft-versus-host disease. J Leukoc Biol 2016; 99: 279-287. [Article] [CrossRef] [PubMed] [Google Scholar]
- Murphy TL, Grajales-Reyes GE, Wu X, et al. Transcriptional control of dendritic cell development. Annu Rev Immunol 2016; 34: 93-119. [Article] [CrossRef] [PubMed] [Google Scholar]
- Durai V, Murphy KM. Functions of murine dendritic cells. Immunity 2016; 45: 719-736. [Article] [CrossRef] [PubMed] [Google Scholar]
- Eisenbarth SC. Dendritic cell subsets in T cell programming: location dictates function. Nat Rev Immunol 2019; 19: 89-103. [Article] [CrossRef] [PubMed] [Google Scholar]
- Izumi G, Nakano H, Nakano K, et al. CD11b+ lung dendritic cells at different stages of maturation induce Th17 or Th2 differentiation. Nat Commun 2021; 12: 5029. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Gargaro M, Scalisi G, Manni G, et al. Indoleamine 2,3-dioxygenase 1 activation in mature cDC1 promotes tolerogenic education of inflammatory cDC2 via metabolic communication. Immunity 2022; 55: 1032-1050.e14. [Article] [CrossRef] [PubMed] [Google Scholar]
- Ardouin L, Luche H, Chelbi R, et al. Broad and largely concordant molecular changes characterize tolerogenic and immunogenic dendritic cell maturation in thymus and periphery. Immunity 2016; 45: 305-318. [Article] [CrossRef] [PubMed] [Google Scholar]
- Barroso A, Mahler JV, Fonseca-Castro PH, et al. Therapeutic induction of tolerogenic dendritic cells via aryl hydrocarbon receptor signaling. Curr Opin Immunol 2021; 70: 33-39. [Article] [CrossRef] [PubMed] [Google Scholar]
- Eickhoff S, Brewitz A, Gerner MY, et al. Robust anti-viral immunity requires multiple distinct T cell-dendritic cell interactions. Cell 2015; 162: 1322-1337. [Article] [CrossRef] [PubMed] [Google Scholar]
- Hor JL, Whitney PG, Zaid A, et al. Spatiotemporally distinct interactions with dendritic cell subsets facilitates CD4+ and CD8+ T cell activation to localized viral infection. Immunity 2015; 43: 554-565. [Article] [CrossRef] [PubMed] [Google Scholar]
- Alcaraz-Serna A, Bustos-Morán E, Fernández-Delgado I, et al. Immune synapse instructs epigenomic and transcriptomic functional reprogramming in dendritic cells. Sci Adv 2021; 7: eabb9965. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Wu R, Murphy KM. DCs at the center of help: Origins and evolution of the three-cell-type hypothesis. J Exp Med 2022; 219: e20211519 [CrossRef] [PubMed] [Google Scholar]
- Ferris ST, Durai V, Wu R, et al. cDC1 prime and are licensed by CD4+ T cells to induce anti-tumour immunity. Nature 2020; 584: 624-629. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Schaupp L, Muth S, Rogell L, et al. Microbiota-induced type I interferons instruct a poised basal state of dendritic cells. Cell 2020; 181: 1080-1096.e19. [Article] [CrossRef] [PubMed] [Google Scholar]
- Zhivaki D, Kagan JC. NLRP3 inflammasomes that induce antitumor immunity. Trends Immunol 2021; 42: 575-589. [Article] [CrossRef] [PubMed] [Google Scholar]
- Zhivaki D, Kagan JC. Innate immune detection of lipid oxidation as a threat assessment strategy. Nat Rev Immunol 2022; 22: 322-330. [Article] [CrossRef] [PubMed] [Google Scholar]
- Zhivaki D, Borriello F, Chow OA, et al. Inflammasomes within hyperactive murine dendritic cells stimulate long-lived T cell-mediated anti-tumor immunity. Cell Rep 2020; 33: 108381. [Article] [CrossRef] [PubMed] [Google Scholar]
- Lee PH, Yamamoto TN, Gurusamy D, et al. Host conditioning with IL-1β improves the antitumor function of adoptively transferred T cells. J Exp Med 2019; 216: 2619-2634. [Article] [CrossRef] [PubMed] [Google Scholar]
- North RJ, Neubauer RH, Huang JJ, et al. Interleukin 1-induced, T cell-mediated regression of immunogenic murine tumors. Requirement for an adequate level of already acquired host concomitant immunity. J Exp Med 1988; 168: 2031-2043. [Article] [CrossRef] [PubMed] [Google Scholar]
- Dixit A, Parnas O, Li B, et al. Perturb-Seq: Dissecting molecular circuits with scalable single-cell RNA profiling of pooled genetic screens. Cell 2016; 167: 1853-1866.e17. [Article] [Google Scholar]
- Grandclaudon M, Perrot-Dockès M, Trichot C, et al. A quantitative multivariate model of human dendritic cell-T helper cell communication. Cell 2019; 179: 432-447.e21. [Article] [CrossRef] [PubMed] [Google Scholar]
- Hoffmann C, Noel F, Grandclaudon M, et al. PD-L1 and ICOSL discriminate human secretory and helper dendritic cells in cancer, allergy and autoimmunity. Nat Commun 2022; 13: 1983. [Article] [CrossRef] [PubMed] [Google Scholar]
- Hilligan KL, Tang SC, Hyde EJ, et al. Dermal IRF4+ dendritic cells and monocytes license CD4+ T helper cells to distinct cytokine profiles. Nat Commun 2020; 11: 5637. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Kvedaraite E, Ginhoux F. Human dendritic cells in cancer. Sci Immunol 2022; 7: eabm9409. [Article] [CrossRef] [PubMed] [Google Scholar]
- Zhang S, Chopin M, Nutt SL. Type 1 conventional dendritic cells: ontogeny, function, and emerging roles in cancer immunotherapy. Trends Immunol 2021; 42: 1113-1127. [Article] [CrossRef] [PubMed] [Google Scholar]
- Jhunjhunwala S, Hammer C, Delamarre L. Antigen presentation in cancer: insights into tumour immunogenicity and immune evasion. Nat Rev Cancer 2021; 21: 298-312. [Article] [CrossRef] [PubMed] [Google Scholar]
- Ghislat G, Cheema AS, Baudoin E, et al. NF-κB-dependent IRF1 activation programs cDC1 dendritic cells to drive antitumor immunity. Sci Immunol 2021; 6: eabg3570. [Article] [CrossRef] [PubMed] [Google Scholar]
- Ruhland MK, Roberts EW, Cai E, et al. Visualizing synaptic transfer of tumor antigens among dendritic cells. Cancer Cell 2020; 37: 786-799.e5. [Article] [CrossRef] [PubMed] [Google Scholar]
- Duong E, Fessenden TB, Lutz E, et al. Type I interferon activates MHC class I-dressed CD11b+ conventional dendritic cells to promote protective anti-tumor CD8+ T cell immunity. Immunity 2022; 55: 308-323.e9. [Article] [CrossRef] [PubMed] [Google Scholar]
- Roberts EW, Broz ML, Binnewies M, et al. Critical role for CD103+/CD141+ dendritic cells bearing CCR7 for tumor antigen trafficking and priming of T cell immunity in melanoma. Cancer Cell 2016; 30: 324-336. [Article] [CrossRef] [PubMed] [Google Scholar]
- Menares E, Gálvez-Cancino F, Cáceres-Morgado P, et al. Tissue-resident memory CD8+ T cells amplify anti-tumor immunity by triggering antigen spreading through dendritic cells. Nat Commun 2019; 10: 4401. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- van den Hout MFCM, Koster BD, Sluijter BJR, et al. Melanoma sequentially suppresses different DC subsets in the sentinel lymph node, affecting disease spread and recurrence. Cancer Immunol Res 2017; 5: 969-977. [Article] [CrossRef] [PubMed] [Google Scholar]
- Möller A, Lobb RJ. The evolving translational potential of small extracellular vesicles in cancer. Nat Rev Cancer 2020; 20: 697-709. [Article] [CrossRef] [PubMed] [Google Scholar]
- Melaiu O, Chierici M, Lucarini V, et al. Cellular and gene signatures of tumor-infiltrating dendritic cells and natural-killer cells predict prognosis of neuroblastoma. Nat Commun 2020; 11: 5992. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Zeng D, Li M, Zhou R, et al. Tumor microenvironment characterization in gastric cancer identifies prognostic and immunotherapeutically relevant gene signatures. Cancer Immunol Res 2019; 7: 737-750. [Article] [CrossRef] [PubMed] [Google Scholar]
- Fuertes MB, Kacha AK, Kline J, et al. Host type I IFN signals are required for antitumor CD8+ T cell responses through CD8α+ dendritic cells. J Exp Med 2011; 208: 2005-2016. [Article] [CrossRef] [PubMed] [Google Scholar]
- Woo SR, Fuertes MB, Corrales L, et al. STING-dependent cytosolic DNA sensing mediates innate immune recognition of immunogenic tumors. Immunity 2014; 41: 830-842. [Article] [CrossRef] [PubMed] [Google Scholar]
- Shi YY, Zheng WX, Yang KT, et al. Intratumoral accumulation of gut microbiota facilitates CD47-based immunotherapy via STING signaling.J Exper Med 2020; 217: e20192282 [CrossRef] [PubMed] [Google Scholar]
- McWhirter SM, Jefferies CA. Nucleic acid sensors as therapeutic targets for human disease. Immunity 2020; 53: 78-97. [Article] [CrossRef] [PubMed] [Google Scholar]
- Nicolai CJ, Wolf N, Chang IC, et al. NK cells mediate clearance of CD8+ T cell-resistant tumors in response to STING agonists. Sci Immunol 2020; 5: eaaz2738 [CrossRef] [PubMed] [Google Scholar]
- Xu MM, Pu Y, Han D, et al. Dendritic cells but not macrophages sense tumor mitochondrial DNA for cross-priming through signal regulatory protein α signaling. Immunity 2017; 47: 363-373.e5. [Article] [CrossRef] [PubMed] [Google Scholar]
- de Mingo Pulido Á, Hänggi K, Celias DP, et al. The inhibitory receptor TIM-3 limits activation of the cGAS-STING pathway in intra-tumoral dendritic cells by suppressing extracellular DNA uptake. Immunity 2021; 54: 1154-1167.e7. [Article] [CrossRef] [PubMed] [Google Scholar]
- He M, Soni B, Schwalie PC, et al. Combinations of Toll-like receptor 8 agonist TL8-506 activate human tumor-derived dendritic cells. J Immunother Cancer 2022; 10: e004268 [CrossRef] [MathSciNet] [Google Scholar]
- Hubert M, Gobbini E, Couillault C, et al. IFN-III is selectively produced by cDC1 and predicts good clinical outcome in breast cancer. Sci Immunol 2020; 5: eaav3942. [Article] [CrossRef] [PubMed] [Google Scholar]
- Martin JD, Cabral H, Stylianopoulos T, et al. Improving cancer immunotherapy using nanomedicines: progress, opportunities and challenges. Nat Rev Clin Oncol 2020; 17: 251-266. [Article] [CrossRef] [PubMed] [Google Scholar]
- Salmon H, Idoyaga J, Rahman A, et al. Expansion and activation of CD103+ dendritic cell progenitors at the tumor site enhances tumor responses to therapeutic PD-L1 and BRAF inhibition. Immunity 2016; 44: 924-938. [Article] [CrossRef] [PubMed] [Google Scholar]
- Hammerich L, Marron TU, Upadhyay R, et al. Systemic clinical tumor regressions and potentiation of PD1 blockade with in situ vaccination. Nat Med 2019; 25: 814-824. [Article] [CrossRef] [PubMed] [Google Scholar]
- Zhang Q, He Y, Luo N, et al. Landscape and dynamics of single immune cells in hepatocellular carcinoma. Cell 2019; 179: 829-845.e20. [Article] [CrossRef] [PubMed] [Google Scholar]
- Maier B, Leader AM, Chen ST, et al. A conserved dendritic-cell regulatory program limits antitumour immunity. Nature 2020; 580: 257-262. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Zilionis R, Engblom C, Pfirschke C, et al. Single-cell transcriptomics of human and mouse lung cancers reveals conserved myeloid populations across individuals and species. Immunity 2019; 50: 1317-1334.e10. [Article] [CrossRef] [PubMed] [Google Scholar]
- Gerhard GM, Bill R, Messemaker M, et al. Tumor-infiltrating dendritic cell states are conserved across solid human cancers. J Exp Med 2021; 218: e20200264. [Article] [CrossRef] [PubMed] [Google Scholar]
- Cheng S, Li Z, Gao R, et al. A pan-cancer single-cell transcriptional atlas of tumor infiltrating myeloid cells. Cell 2021; 184: 792-809.e23. [Article] [CrossRef] [PubMed] [Google Scholar]
- Combes AJ, Samad B, Tsui J, et al. Discovering dominant tumor immune archetypes in a pan-cancer census. Cell 2022; 185: 184-203.e19. [Article] [CrossRef] [PubMed] [Google Scholar]
- Smalley I, Chen Z, Phadke M, et al. Single-cell characterization of the immune microenvironment of melanoma brain and leptomeningeal metastases. Clin Cancer Res 2021; 27: 4109-4125. [Article] [CrossRef] [PubMed] [Google Scholar]
- Ji AL, Rubin AJ, Thrane K, et al. Multimodal analysis of composition and spatial architecture in human squamous cell carcinoma (vol 182, pg 497, 2020). Cell 2020; 182: 1661–1662 [CrossRef] [PubMed] [Google Scholar]
- Leruste A, Tosello J, Ramos RN, et al. Clonally expanded T cells reveal immunogenicity of rhabdoid tumors. Cancer Cell 2019; 36: 597-612.e8. [Article] [CrossRef] [PubMed] [Google Scholar]
- Steele NG, Carpenter ES, Kemp SB, et al. Multimodal mapping of the tumor and peripheral blood immune landscape in human pancreatic cancer. Nat Cancer 2020; 1: 1097-1112. [Article] [CrossRef] [PubMed] [Google Scholar]
- Hornburg M, Desbois M, Lu S, et al. Single-cell dissection of cellular components and interactions shaping the tumor immune phenotypes in ovarian cancer. Cancer Cell 2021; 39: 928-944.e6. [Article] [CrossRef] [PubMed] [Google Scholar]
- Cohen M, Giladi A, Barboy O, et al. The interaction of CD4+ helper T cells with dendritic cells shapes the tumor microenvironment and immune checkpoint blockade response. Nat Cancer 2022; 3: 303-317. [Article] [CrossRef] [PubMed] [Google Scholar]
- Wu FY, Fan J, He YY, et al. Single-cell profiling of tumor heterogeneity and the microenvironment in advanced non-small cell lung cancer. Nat Commun 2021; 12: 2540 [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Leader AM, Grout JA, Maier BB, et al. Single-cell analysis of human non-small cell lung cancer lesions refines tumor classification and patient stratification. Cancer Cell 2021; 39: 1594-1609.e12. [Article] [CrossRef] [PubMed] [Google Scholar]
- Peng WS, Zhou X, Yan WB, et al. Dissecting the heterogeneity of the microenvironment in primary and recurrent nasopharyngeal carcinomas using single-cell RNA sequencing. OncoImmunology 2022; 11: 2026583. [Article] [CrossRef] [PubMed] [Google Scholar]
- Chen YP, Yin JH, Li WF, et al. Single-cell transcriptomics reveals regulators underlying immune cell diversity and immune subtypes associated with prognosis in nasopharyngeal carcinoma. Cell Res 2020; 30: 1024-1042. [Article] [CrossRef] [PubMed] [Google Scholar]
- Liu Y, He S, Wang XL, et al. Tumour heterogeneity and intercellular networks of nasopharyngeal carcinoma at single cell resolution. Nat Commun 2021; 12: 741 [CrossRef] [PubMed] [Google Scholar]
- Sun Y, Wu L, Zhong Y, et al. Single-cell landscape of the ecosystem in early-relapse hepatocellular carcinoma. Cell 2021; 184: 404-421.e16. [Article] [CrossRef] [PubMed] [Google Scholar]
- Mair F, Erickson JR, Frutoso M, et al. Extricating human tumour immune alterations from tissue inflammation. Nature 2022; 605: 728-735. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Cillo AR, Kürten CHL, Tabib T, et al. Immune landscape of viral- and carcinogen-driven head and neck cancer. Immunity 2020; 52: 183-199.e9. [Article] [CrossRef] [PubMed] [Google Scholar]
- Zheng Y, Chen Z, Han Y, et al. Immune suppressive landscape in the human esophageal squamous cell carcinoma microenvironment. Nat Commun 2020; 11: 6268. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Dinh HQ, Pan F, Wang G, et al. Integrated single-cell transcriptome analysis reveals heterogeneity of esophageal squamous cell carcinoma microenvironment. Nat Commun 2021; 12: 7335. [Article] [CrossRef] [PubMed] [Google Scholar]
- Liu Y, Zhang Q, Xing B, et al. Immune phenotypic linkage between colorectal cancer and liver metastasis. Cancer Cell 2022; 40: 424-437.e5. [Article] [CrossRef] [PubMed] [Google Scholar]
- Zhang L, Li Z, Skrzypczynska KM, et al. Single-cell analyses inform mechanisms of myeloid-targeted therapies in colon cancer. Cell 2020; 181: 442-459.e29. [Article] [CrossRef] [PubMed] [Google Scholar]
- Wu SZ, Al-Eryani G, Roden DL, et al. A single-cell and spatially resolved atlas of human breast cancers. Nat Genet 2021; 53: 1334-1347. [Article] [CrossRef] [PubMed] [Google Scholar]
- Bassez A, Vos H, Van Dyck L, et al. A single-cell map of intratumoral changes during anti-PD1 treatment of patients with breast cancer. Nat Med 2021; 27: 820-832. [Article] [CrossRef] [PubMed] [Google Scholar]
- Zhang Y, Chen H, Mo H, et al. Single-cell analyses reveal key immune cell subsets associated with response to PD-L1 blockade in triple-negative breast cancer. Cancer Cell 2021; 39: 1578-1593.e8. [Article] [CrossRef] [PubMed] [Google Scholar]
- Chen Z, Zhou L, Liu L, et al. Single-cell RNA sequencing highlights the role of inflammatory cancer-associated fibroblasts in bladder urothelial carcinoma. Nat Commun 2020; 11: 5077. [Article] [CrossRef] [MathSciNet] [PubMed] [Google Scholar]
- Zhou Y, Yang D, Yang Q, et al. Single-cell RNA landscape of intratumoral heterogeneity and immunosuppressive microenvironment in advanced osteosarcoma. Nat Commun 2020; 11: 6322. [Article] [CrossRef] [PubMed] [Google Scholar]
- Tan Y, Flynn WF, Sivajothi S, et al. Single-cell analysis of endometriosis reveals a coordinated transcriptional programme driving immunotolerance and angiogenesis across eutopic and ectopic tissues. Nat Cell Biol 2022; 24: 1306-1318. [Article] [CrossRef] [PubMed] [Google Scholar]
- Liu Y, Wang H, Taylor M, et al. Classification of human chronic inflammatory skin disease based on single-cell immune profiling. Sci Immunol 2022; 7: eabl9165. [Article] [CrossRef] [PubMed] [Google Scholar]
- Hildreth AD, Ma F, Wong YY, et al. Single-cell sequencing of human white adipose tissue identifies new cell states in health and obesity. Nat Immunol 2021; 22: 639-653. [Article] [CrossRef] [PubMed] [Google Scholar]
- Mulder K, Patel AA, Kong WT, et al. Cross-tissue single-cell landscape of human monocytes and macrophages in health and disease. Immunity 2021; 54: 1883-1900.e5. [Article] [CrossRef] [PubMed] [Google Scholar]
- Nakamizo S, Dutertre CA, Khalilnezhad A, et al. Single-cell analysis of human skin identifies CD14+ type 3 dendritic cells co-producing IL1B and IL23A in psoriasis. J Exp Med 2021; 218: e20202345. [Article] [CrossRef] [PubMed] [Google Scholar]
- Reynolds G, Vegh P, Fletcher J, et al. Developmental cell programs are co-opted in inflammatory skin disease. Science 2021; 371: 364. [Article] [NASA ADS] [CrossRef] [Google Scholar]
- Park JE, Botting RA, Conde CD, et al. A cell atlas of human thymic development defines t cell repertoire formation. Science 2020; 367: 868 [NASA ADS] [Google Scholar]
- Dutertre CA, Becht E, Irac SE, et al. Single-cell analysis of human mononuclear phagocytes reveals subset-defining markers and identifies circulating inflammatory dendritic cells. Immunity 2019; 51: 573-589.e8. [Article] [CrossRef] [PubMed] [Google Scholar]
- Cytlak U, Resteu A, Pagan S, et al. Differential IRF8 transcription factor requirement defines two pathways of dendritic cell development in humans. Immunity 2020; 53: 353-370.e8. [Article] [CrossRef] [PubMed] [Google Scholar]
- Di Pilato M, Kfuri-Rubens R, Pruessmann JN, et al. CXCR6 positions cytotoxic T cells to receive critical survival signals in the tumor microenvironment. Cell 2021; 184: 4512-4530.e22. [Article] [CrossRef] [PubMed] [Google Scholar]
- Kreatsoulas D, Bolyard C, Wu BX, et al. Translational landscape of glioblastoma immunotherapy for physicians: guiding clinical practice with basic scientific evidence. J Hematol Oncol 2022; 15: 80. [Article] [CrossRef] [PubMed] [Google Scholar]
- Peng Q, Qiu X, Zhang Z, et al. PD-L1 on dendritic cells attenuates T cell activation and regulates response to immune checkpoint blockade. Nat Commun 2020; 11: 4835. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Dähling S, Mansilla AM, Knöpper K, et al. Type 1 conventional dendritic cells maintain and guide the differentiation of precursors of exhausted T cells in distinct cellular niches. Immunity 2022; 55: 656-670.e8. [Article] [CrossRef] [PubMed] [Google Scholar]
- Schenkel JM, Herbst RH, Canner D, et al. Conventional type I dendritic cells maintain a reservoir of proliferative tumor-antigen specific TCF-1+ CD8+ T cells in tumor-draining lymph nodes. Immunity 2021; 54: 2338-2353.e6. [Article] [CrossRef] [PubMed] [Google Scholar]
- Sautès-Fridman C, Petitprez F, Calderaro J, et al. Tertiary lymphoid structures in the era of cancer immunotherapy. Nat Rev Cancer 2019; 19: 307-325. [Article] [CrossRef] [PubMed] [Google Scholar]
- Bruni D, Angell HK, Galon J. The immune contexture and Immunoscore in cancer prognosis and therapeutic efficacy. Nat Rev Cancer 2020; 20: 662-680. [Article] [CrossRef] [PubMed] [Google Scholar]
- Bai X, Zhou Y, Yokota Y, et al. Adaptive antitumor immune response stimulated by bio-nanoparticle based vaccine and checkpoint blockade. J Exp Clin Cancer Res 2022; 41: 132. [Article] [CrossRef] [PubMed] [Google Scholar]
- Vuong L, Kotecha RR, Voss MH, et al. Tumor microenvironment dynamics in clear-cell renal cell carcinoma. Cancer Discov 2019; 9: 1349-1357. [Article] [CrossRef] [PubMed] [Google Scholar]
- Giraldo NA, Becht E, Pagès F, et al. Orchestration and prognostic significance of immune checkpoints in the microenvironment of primary and metastatic renal cell cancer. Clin Cancer Res 2015; 21: 3031-3040. [Article] [CrossRef] [PubMed] [Google Scholar]
- van Hooren L, Vaccaro A, Ramachandran M, et al. Agonistic CD40 therapy induces tertiary lymphoid structures but impairs responses to checkpoint blockade in glioma. Nat Commun 2021; 12: 4127. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Chelvanambi M, Fecek RJ, Taylor JL, et al. STING agonist-based treatment promotes vascular normalization and tertiary lymphoid structure formation in the therapeutic melanoma microenvironment. J Immunother Cancer 2021; 9: e001906. [Article] [CrossRef] [PubMed] [Google Scholar]
- Fridman WH, Meylan M, Petitprez F, et al. B cells and tertiary lymphoid structures as determinants of tumour immune contexture and clinical outcome. Nat Rev Clin Oncol 2022; 19: 441-457. [Article] [CrossRef] [PubMed] [Google Scholar]
- Kalergis AM, Ravetch JV. Inducing tumor immunity through the selective engagement of activating Fcγ receptors on dendritic cells. J Exp Med 2002; 195: 1653-1659. [Article] [CrossRef] [PubMed] [Google Scholar]
- Dieu-Nosjean MC, Antoine M, Danel C, et al. Long-term survival for patients with non-small-cell lung cancer with intratumoral lymphoid structures. J Clin Oncol 2008; 26: 4410-4417. [Article] [CrossRef] [PubMed] [Google Scholar]
- Clark DJ, Dhanasekaran SM, Petralia F, et al. Integrated proteogenomic characterization of clear cell renal cell carcinoma. Cell 2019; 179: 964-983.e31. [Article] [CrossRef] [PubMed] [Google Scholar]
- Meyer MA, Baer JM, Knolhoff BL, et al. Breast and pancreatic cancer interrupt IRF8-dependent dendritic cell development to overcome immune surveillance. Nat Commun 2018; 9: 1250. [Article] [CrossRef] [PubMed] [Google Scholar]
- Devalaraja S, To TKJ, Folkert IW, et al. Tumor-derived retinoic acid regulates intratumoral monocyte differentiation to promote immune suppression. Cell 2020; 180: 1098-1114.e16. [Article] [CrossRef] [PubMed] [Google Scholar]
- Hegde S, Krisnawan VE, Herzog BH, et al. Dendritic cell paucity leads to dysfunctional immune surveillance in pancreatic cancer. Cancer Cell 2020; 37: 289-307.e9. [Article] [Google Scholar]
- Medina BD, Liu M, Vitiello GA, et al. Oncogenic kinase inhibition limits Batf3-dependent dendritic cell development and antitumor immunity. J Exp Med 2019; 216: 1359-1376. [Article] [CrossRef] [PubMed] [Google Scholar]
- Giovanelli P, Sandoval TA, Cubillos-Ruiz JR. Dendritic cell metabolism and function in tumors. Trends Immunol 2019; 40: 699-718. [Article] [CrossRef] [PubMed] [Google Scholar]
- Böttcher JP, Bonavita E, Chakravarty P, et al. NK cells stimulate recruitment of cDC1 into the tumor microenvironment promoting cancer immune control. Cell 2018; 172: 1022-1037.e14. [Article] [CrossRef] [PubMed] [Google Scholar]
- Hayashi K, Nikolos F, Lee YC, et al. Tipping the immunostimulatory and inhibitory DAMP balance to harness immunogenic cell death. Nat Commun 2020; 11: 6299. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Bai X, Wong CC, Pan Y, et al. Loss of YTHDF1 in gastric tumors restores sensitivity to antitumor immunity by recruiting mature dendritic cells. J Immunother Cancer 2022; 10: e003663. [Article] [CrossRef] [PubMed] [Google Scholar]
- Han D, Liu J, Chen C, et al. Anti-tumour immunity controlled through mRNA m6A methylation and YTHDF1 in dendritic cells. Nature 2019; 566: 270-274. [Article] [NASA ADS] [CrossRef] [PubMed] [Google Scholar]
- Galon J, Bruni D. Approaches to treat immune hot, altered and cold tumours with combination immunotherapies. Nat Rev Drug Discov 2019; 18: 197-218. [Article] [CrossRef] [PubMed] [Google Scholar]
- Munson PV, Adamik J, Butterfield LH. Immunomodulatory impact of α-fetoprotein. Trends Immunol 2022; 43: 438-448. [Article] [CrossRef] [PubMed] [Google Scholar]
- Moller SH, Wang LM, Ho PC. Metabolic programming in dendritic cells tailors immune responses and homeostasis. Cell Mol Immuno 2022; 19: 370–383 [CrossRef] [PubMed] [Google Scholar]
- Colgan SP, Furuta GT, Taylor CT. Hypoxia and innate immunity: Keeping up with the HIFsters. Annu Rev Immunol 2020; 38: 341-363. [Article] [CrossRef] [PubMed] [Google Scholar]
- Suthen S, Lim CJ, Nguyen PHD, et al. Hypoxia-driven immunosuppression by Treg and type-2 conventional dendritic cells in HCC. Hepatology 2022; 76: 1329-1344. [Article] [CrossRef] [PubMed] [Google Scholar]
- McGettrick AF, O’Neill LAJ. The role of HIF in immunity and inflammation. Cell Metab 2020; 32: 524-536. [Article] [CrossRef] [PubMed] [Google Scholar]
- Song M, Cubillos-Ruiz JR. Endoplasmic reticulum stress responses in intratumoral immune cells: implications for cancer immunotherapy. Trends Immunol 2019; 40: 128-141. [Article] [CrossRef] [PubMed] [Google Scholar]
- Hinshaw DC, Shevde LA. The tumor microenvironment innately modulates cancer progression. Cancer Res 2019; 79: 4557-4566. [Article] [CrossRef] [Google Scholar]
- Allard B, Allard D, Buisseret L, et al. The adenosine pathway in immuno-oncology. Nat Rev Clin Oncol 2020; 17: 611-629. [Article] [CrossRef] [PubMed] [Google Scholar]
- Friedmann Angeli JP, Krysko DV, Conrad M. Ferroptosis at the crossroads of cancer-acquired drug resistance and immune evasion. Nat Rev Cancer 2019; 19: 405-414. [Article] [CrossRef] [PubMed] [Google Scholar]
- Garner H, de Visser KE. Immune crosstalk in cancer progression and metastatic spread: a complex conversation. Nat Rev Immunol 2020; 20: 483-497. [Article] [CrossRef] [PubMed] [Google Scholar]
- Bader JE, Voss K, Rathmell JC. Targeting metabolism to improve the tumor microenvironment for cancer immunotherapy. Mol Cell 2020; 78: 1019-1033. [Article] [CrossRef] [PubMed] [Google Scholar]
- Yuan X, Duan Y, Xiao Y, et al. Vitamin E enhances cancer immunotherapy by reinvigorating dendritic cells via targeting checkpoint SHP1. Cancer Discov 2022; 12: 1742-1759. [Article] [CrossRef] [MathSciNet] [PubMed] [Google Scholar]
- Uribe-Herranz M, Rafail S, Beghi S, et al. Gut microbiota modulate dendritic cell antigen presentation and radiotherapy-induced antitumor immune response. J Clin Invest 2020; 130: 466-479. [Article] [CrossRef] [Google Scholar]
- Huang TX, Tan XY, Huang HS, et al. Targeting cancer-associated fibroblast-secreted WNT2 restores dendritic cell-mediated antitumour immunity. Gut 2022; 71: 333-344. [Article] [CrossRef] [PubMed] [Google Scholar]
- Ebina-Shibuya R, Leonard WJ. Role of thymic stromal lymphopoietin in allergy and beyond. Nat Rev Immunol, 2023; 23: 24–37 [CrossRef] [PubMed] [Google Scholar]
- Santana-Magal N, Farhat-Younis L, Gutwillig A, et al. Melanoma-secreted lysosomes trigger monocyte-derived dendritic cell apoptosis and limit cancer immunotherapy. Cancer Res 2020; 80: 1942-1956. [Article] [CrossRef] [PubMed] [Google Scholar]
- Ozga AJ, Chow MT, Luster AD. Chemokines and the immune response to cancer. Immunity 2021; 54: 859-874. [Article] [CrossRef] [PubMed] [Google Scholar]
- Barry KC, Hsu J, Broz ML, et al. A natural killer-dendritic cell axis defines checkpoint therapy-responsive tumor microenvironments. Nat Med 2018; 24: 1178-1191. [Article] [CrossRef] [PubMed] [Google Scholar]
- Binnewies M, Mujal AM, Pollack JL, et al. Unleashing type-2 dendritic cells to drive protective antitumor CD4+ T cell immunity. Cell 2019; 177: 556-571.e16. [Article] [CrossRef] [PubMed] [Google Scholar]
- Batlle E, Massagué J. Transforming growth factor-β signaling in immunity and cancer. Immunity 2019; 50: 924-940. [Article] [CrossRef] [PubMed] [Google Scholar]
- Marangoni F, Zhakyp A, Corsini M, et al. Expansion of tumor-associated Treg cells upon disruption of a CTLA-4-dependent feedback loop. Cell 2021; 184: 3998-4015.e19. [Article] [CrossRef] [PubMed] [Google Scholar]
- Leinwand J, Miller G. Regulation and modulation of antitumor immunity in pancreatic cancer. Nat Immunol 2020; 21: 1152-1159. [Article] [CrossRef] [PubMed] [Google Scholar]
- Bullock TNJ. CD40 stimulation as a molecular adjuvant for cancer vaccines and other immunotherapies. Cell Mol Immunol 2022; 19: 14-22. [Article] [CrossRef] [PubMed] [Google Scholar]
- Garris CS, Wong JL, Ravetch JV, et al. Dendritic cell targeting with Fc-enhanced CD40 antibody agonists induces durable antitumor immunity in humanized mouse models of bladder cancer. Sci Transl Med 2021; 13: eabd1346. [Article] [CrossRef] [PubMed] [Google Scholar]
- Burrack AL, Schmiechen ZC, Patterson MT, et al. Distinct myeloid antigen-presenting cells dictate differential fates of tumor-specific CD8+ T cells in pancreatic cancer. JCI Insight 2022; 7: e151593. [Article] [CrossRef] [PubMed] [Google Scholar]
- Dammeijer F, van Gulijk M, Mulder EE, et al. The PD-1/PD-L1-checkpoint restrains T cell immunity in tumor-draining lymph nodes. Cancer Cell 2020; 38: 685-700.e8. [Article] [CrossRef] [PubMed] [Google Scholar]
- Nirmal AJ, Maliga Z, Vallius T, et al. The spatial landscape of progression and immunoediting in primary melanoma at single-cell resolution. Cancer Discov 2022; 12: 1518-1541. [Article] [CrossRef] [PubMed] [Google Scholar]
- Oh SA, Wu DC, Cheung J, et al. PD-L1 expression by dendritic cells is a key regulator of T-cell immunity in cancer. Nat Cancer 2020; 1: 681-691. [Article] [CrossRef] [PubMed] [Google Scholar]
- Zhao Y, Lee CK, Lin CH, et al. PD-L1:CD80 cis-heterodimer triggers the co-stimulatory receptor CD28 while repressing the inhibitory PD-1 and CTLA-4 pathways. Immunity 2019; 51: 1059-1073.e9. [Article] [CrossRef] [PubMed] [Google Scholar]
- Mayoux M, Roller A, Pulko V, et al. Dendritic cells dictate responses to PD-L1 blockade cancer immunotherapy. Sci Transl Med 2020; 12: eaav7431. [Article] [CrossRef] [PubMed] [Google Scholar]
- Topalian SL, Taube JM, Pardoll DM. Neoadjuvant checkpoint blockade for cancer immunotherapy. Science 2020; 367: 525. [Article] [NASA ADS] [CrossRef] [Google Scholar]
- Garris CS, Arlauckas SP, Kohler RH, et al. Successful anti-PD-1 cancer immunotherapy requires T cell-dendritic cell crosstalk involving the cytokines IFN-γ and IL-12. Immunity 2018; 49: 1148-1161.e7. [Article] [CrossRef] [PubMed] [Google Scholar]
- Chow MT, Ozga AJ, Servis RL, et al. Intratumoral activity of the CXCR3 chemokine system is required for the efficacy of anti-PD-1 therapy. Immunity 2019; 50: 1498-1512.e5. [Article] [CrossRef] [PubMed] [Google Scholar]
- Chen L, Sun R, Xu J, et al. Tumor-derived IL33 promotes tissue-resident CD8+ T cells and is required for checkpoint blockade tumor immunotherapy. Cancer Immunol Res 2020; 8: 1381-1392. [Article] [CrossRef] [PubMed] [Google Scholar]
- de Mingo Pulido Á, Gardner A, Hiebler S, et al. TIM-3 regulates CD103+ dendritic cell function and response to chemotherapy in breast cancer. Cancer Cell 2018; 33: 60-74.e6. [Article] [CrossRef] [PubMed] [Google Scholar]
- Gardner A, de Mingo Pulido Á, Hänggi K, et al. TIM-3 blockade enhances IL-12-dependent antitumor immunity by promoting CD8+ T cell and XCR1+ dendritic cell spatial co-localization. J Immunother Cancer 2022; 10: e003571. [Article] [CrossRef] [PubMed] [Google Scholar]
- Feng M, Jiang W, Kim BYS, et al. Phagocytosis checkpoints as new targets for cancer immunotherapy. Nat Rev Cancer 2019; 19: 568-586. [Article] [CrossRef] [PubMed] [Google Scholar]
- Dougan M, Dranoff G, Dougan SK. Cancer immunotherapy: beyond checkpoint blockade. Annu Rev Cancer Biol 2019; 3: 55-75. [Article] [CrossRef] [PubMed] [Google Scholar]
- Liu Q, Wen W, Tang L, et al. Inhibition of SIRPα in dendritic cells potentiates potent antitumor immunity. OncoImmunology 2016; 5: e1183850. [Article] [CrossRef] [PubMed] [Google Scholar]
- Wang S, Wu Q, Chen T, et al. Blocking CD47 promotes antitumour immunity through CD103+ dendritic cell-NK cell axis in murine hepatocellular carcinoma model. J Hepatol 2022; 77: 467-478. [Article] [CrossRef] [PubMed] [Google Scholar]
- Gauttier V, Pengam S, Durand J, et al. Selective SIRPα blockade reverses tumor T cell exclusion and overcomes cancer immunotherapy resistance. J Clin Invest 2020; 130: 6109-6123. [Article] [CrossRef] [PubMed] [Google Scholar]
- Kepp O, Marabelle A, Zitvogel L, et al. Oncolysis without viruses—inducing systemic anticancer immune responses with local therapies. Nat Rev Clin Oncol 2020; 17: 49-64. [Article] [CrossRef] [PubMed] [Google Scholar]
- Rodriguez-Ruiz ME, Vitale I, Harrington KJ, et al. Immunological impact of cell death signaling driven by radiation on the tumor microenvironment. Nat Immunol 2020; 21: 120-134. [Article] [CrossRef] [PubMed] [Google Scholar]
- Zheng W, Ranoa DRE, Huang X, et al. RIG-I-like receptor LGP2 is required for tumor control by radiotherapy. Cancer Res 2020; 80: 5633-5641. [Article] [CrossRef] [PubMed] [Google Scholar]
- Herrera FG, Ronet C, Ochoa de Olza M, et al. Low-dose radiotherapy reverses tumor immune desertification and resistance to immunotherapy. Cancer Discov 2022; 12: 108-133. [Article] [CrossRef] [PubMed] [Google Scholar]
- Pilones KA, Charpentier M, Garcia-Martinez E, et al. Radiotherapy cooperates with IL15 to induce antitumor immune responses. Cancer Immunol Res 2020; 8: 1054-1063. [Article] [CrossRef] [PubMed] [Google Scholar]
- Mender I, Zhang A, Ren Z, et al. Telomere stress potentiates STING-dependent anti-tumor immunity. Cancer Cell 2020; 38: 400-411.e6. [Article] [CrossRef] [PubMed] [Google Scholar]
- Haas L, Elewaut A, Gerard CL, et al. Acquired resistance to anti-MAPK targeted therapy confers an immune-evasive tumor microenvironment and cross-resistance to immunotherapy in melanoma. Nat Cancer 2021; 2: 693-708. [Article] [CrossRef] [PubMed] [Google Scholar]
- Galluzzi L, Humeau J, Buqué A, et al. Immunostimulation with chemotherapy in the era of immune checkpoint inhibitors. Nat Rev Clin Oncol 2020; 17: 725-741. [Article] [CrossRef] [PubMed] [Google Scholar]
All Tables
Human LAMP3+ DCs described in the single cell transcriptomics of health and diseases
All Figures
|
Figure 1 The stepwise model of DC maturation. The semi-mature DCs are “ready” to receive further stimulations to become fully-mature DCs and to perform diverse functions. |
| In the text | |
|
Figure 2 The potential functional heterogeneity of LAMP3+ DCs. (A) cDC1-deriverd LAMP3+ DCs mainly produce CXCL16 and IL-15, which promote the recruitment and maintenance of CD8 T cells in the TME and therefore enhance the antitumor immune effects. cDC2-deriverd LAMP3+ DCs highly express CCL17, CCL22 and IDO1, which promote the recruitment and differentiation of Treg in the TME and suppress the antitumor immune response. (B) LAMP3+ DCs within TLSs actively recruit and interact with T cells to elicit neoantigen-specific T cell activation, or Treg production. LAMP3+ DCs outside of TLSs exhibit impaired antigen-presenting ability and may be involved in immunosuppression. |
| In the text | |
|
Figure 3 Mechanisms regulating the tumor-infiltrating DC maturation. Tumor cells, CAFs, and tumor-infiltrating microbes inhibit the maturation of tumor-infiltrating DCs by regulating metabolism, etc. The effector T cells and NK cells in the tumor microenvironment promote the generation of LAMP3+ DCs by producing chemokines and IFNs, and immunosuppressive cells such as Tregs inhibit the generation of LAMP3+ DCs by producing IL-10 and TGF-β. The efficacy of radiotherapy and other treatments also depends on the induction of LAMP3+ DCs. The efficacy of radiotherapy, chemotherapy and immunotherapy is also dependent on their induction of LAMP3+ DCs. |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.